
Si potrebbe iniziare citando un famoso film Tanto tempo fa, in una galassia lontana lontana, ma in realtà la nostra storia, davvero particolare e anomala inizia, su scala geologica, neanche tanto tempo fa e in un luogo non tanto lontano.
 Le nostre origini, quelle del genere Homo ed in particolare di noi Homo sapiens, iniziano circa 6 milioni di anni fa in Africa.
Le nostre origini, quelle del genere Homo ed in particolare di noi Homo sapiens, iniziano circa 6 milioni di anni fa in Africa.
Considerata la rarità dei reperti e le difficoltà di eseguire esami sul DNA, ricostruire tutti i passi evolutivi che hanno portato allo sviluppo di Homo sapiens non è compito semplice. I paleoantropologi spesso si trovano a classificare un nostro antenato o un nostro cugino avendo a disposizione solo una falange o un dente. Nonostante queste difficoltà è stato ipotizzato un processo evolutivo che illustreremo.
Prima di arrivare a parlare della storia di Homo riepiloghiamo ciò che è avvenuto ancora prima secondo le ultime teorie.
 Circa 13 milioni di anni fa, dalla famiglia Hominidae si separarono due gruppi che formarono i due rami evolutivi principali (sottofamiglie):
Circa 13 milioni di anni fa, dalla famiglia Hominidae si separarono due gruppi che formarono i due rami evolutivi principali (sottofamiglie):
1) sottofamiglia Homininae (Ominine)
2) sottofamiglia Ponginae
Quest’ultima porterà all’attuale Pongo (comunemente chiamati oranghi).
 Facciamo un salto di 5 milioni di anni e quindi circa 8 milioni di anni fa la sottofamiglia Homininae (Ominine) si separò ulteriormente in due rami (tribù):
Facciamo un salto di 5 milioni di anni e quindi circa 8 milioni di anni fa la sottofamiglia Homininae (Ominine) si separò ulteriormente in due rami (tribù):
1) tribù Hominini (Ominini)
2) tribù Gorillini
Quest’ultima porterà all’attuale genere Gorilla.
 Si fa un altro salto questa volta di appena 2 milioni di anni, quindi circa 6 milioni di anni fa la tribù Hominini si separò ulteriormente in due rami (sottotribù):
Si fa un altro salto questa volta di appena 2 milioni di anni, quindi circa 6 milioni di anni fa la tribù Hominini si separò ulteriormente in due rami (sottotribù):
1) sottotribù Hominina
2) sottotribù Panina
Quest’ultima porterà all’attuale genere Pan che comprende scimpanzé e bonobo.
Qui una tabella riassuntiva:
Scimpanzé, bonobo, gorilla, oranghi e gibboni sono considerati scimmie antropomorfe (cioè simili all’uomo). Tulle le altre scimmie, derivanti dall’altro ramo della superfamiglia Hominoidea, invece sono non antropomorfe.
A questo punto dell’evoluzione iniziamo a trovare qualcosa che inizia ad assomigliarci, in cui riusciamo vagamente a riconoscere qualcosa di familiare.
Dopo la separazione dal progenitore comune circa 6 milioni di anni fa, i generi si susseguirono temporalmente formando un ampio cespuglio, o ramo frondoso come dicono alcuni. Ad oggi sono stati classificati i seguenti generi e per alcuni di essi è stato possibile suddividerli in specie:
- Orrorin tugenensis chiamato anche Millenium Man perché scoperto nell’anno 2000 (circa 6 milioni di anni fa)
- Ardipithecus (circa 5,7-4,4 milioni di anni fa)
- Australopithecus (circa 4,2 milioni di anni fa-?)
- Kenyanthropus platyops (per ora un solo fossile datato tra 3,5-3,2 milioni di anni fa)
- Paranthropus (circa 2,6-1,2 milioni di anni fa)
- Homo (? – attualmente vivente con l’unica specie superstite Homo sapiens, noi)
Per alcuni (Orrorin, Ardipithecus e Kenyantropus) purtroppo sono stati trovati solo pochissimi esemplari fossili o addirittura un solo esemplare fossile (Orrorin e Kenyanthropus).
Di questi generi il più famoso è sicuramente quello delle cosiddette australopitecine che comprende i due generi: Australopithecus e Paranthropus. Gli australopitechi propriamente detti avevano una struttura sia fisica che mascellare-mandibolare (e relativi denti) più gracile, mentre i parantropi avevano una struttura più robusta, con denti più grandi e adatti a masticare cibi più coriacei.
Di tutti i fossili di ominini finora ritrovati sicuramente il più famoso è Lucy. Fu scoperta in Etiopia nel 1974 e consiste in centinaia di frammenti di ossa che rappresentano il 40% dello scheletro di un esemplare femmina di Australopithecus afarensis. Ha costituito una svolta storica in campo paleontologico.

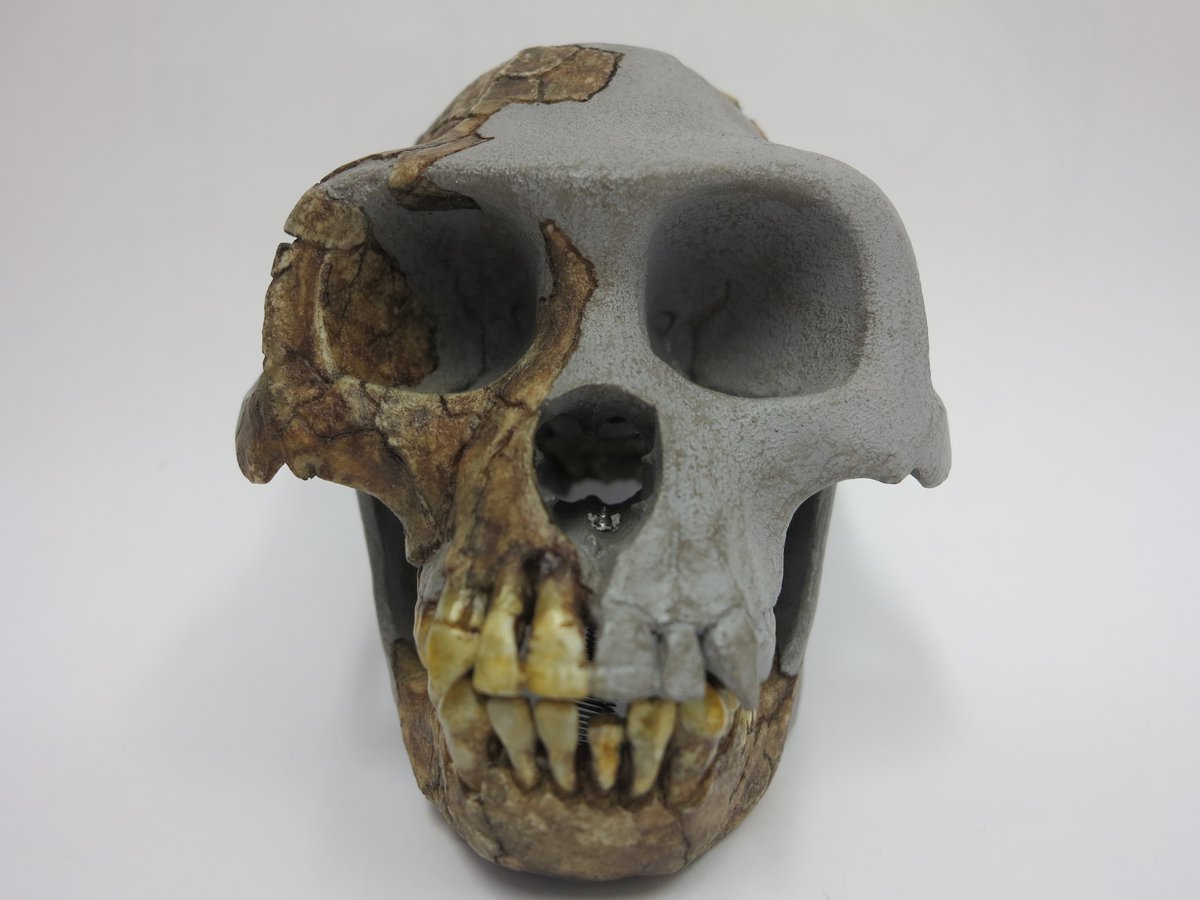
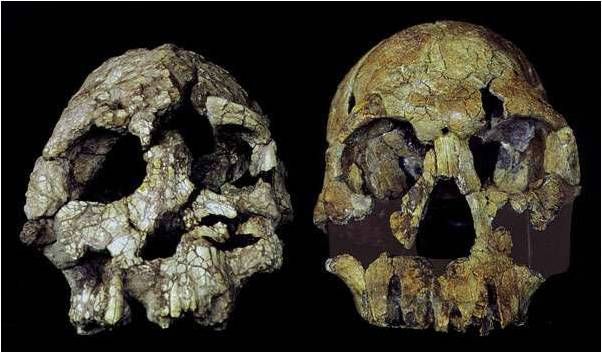


Finalmente siamo arrivati al genere Homo e, prima di fare alcune interessanti considerazioni sulle peculiarità della specie Homo sapiens, vediamo l’attuale suddivisione del genere in taxa (specie):
- Homo habilis (circa 2,4 – 1,4 mln di anni fa)
- Homo rudolfensis (unico esemplare datato circa 1,9 mln di anni fa, KNM-ER 1470)
- Homo georgicus (circa 1,8 mln di anni fa)
- Homo egaster/erectus (circa 1,8 – forse 100.000 anni fa)
- Homo antecessor (circa 1,2 mln – 800.000 anni fa)
- Homo heidelbergensis (circa 600.000 – 100.000 anni fa)
- Homo neanderthalensis (circa 200.000 – 40.000 anni fa)
- Homo denisova (circa 70.000 – 40.000 anni fa)
- Homo naledi (circa 336.000 – 235.000 anni fa)
- Homo floresiensis (190.000 – 54.000 anni fa, forse fino a 12.000 anni fa)
- Homo sapiens (circa 200.000 anni fa – oggi).
Alla luce della mera classificazione, ci sono diverse considerazioni e precisazioni da fare.
Innanzitutto, per quanto concerne le datazioni, queste si riferiscono esclusivamente agli esemplari recuperati dal registro fossile. Ciò significa che vanno considerate in senso relativo (al registro fossile) e non assoluto (alla reale emersione/comparsa della specie e/o del genere in questione).
 Ad esempio, se trovo un esemplare ”X” e lo dato, la datazione è relativa solo a quando è vissuto o, per meglio dire, a quando è morto quell’individuo al quale appartenevano i resti rinvenuti. Ma la specie, o il genere, può essere comparsa anche molto più anticamente. Il registro fossile purtroppo è piuttosto lacunoso e povero in quanto la fossilizzazione è un processo molto raro. Non tutti i terreni sono adatti alla fossilizzazione e non tutti gli organismi hanno avuto la possibilità di fossilizzarsi e, tantomeno, a conservarsi fino ad oggi, per cui le datazioni vanno sempre prese in senso relativo, non in senso assoluto.
Ad esempio, se trovo un esemplare ”X” e lo dato, la datazione è relativa solo a quando è vissuto o, per meglio dire, a quando è morto quell’individuo al quale appartenevano i resti rinvenuti. Ma la specie, o il genere, può essere comparsa anche molto più anticamente. Il registro fossile purtroppo è piuttosto lacunoso e povero in quanto la fossilizzazione è un processo molto raro. Non tutti i terreni sono adatti alla fossilizzazione e non tutti gli organismi hanno avuto la possibilità di fossilizzarsi e, tantomeno, a conservarsi fino ad oggi, per cui le datazioni vanno sempre prese in senso relativo, non in senso assoluto.
Per capire la complessità che ci si trova davanti nell’attribuire una datazione ad una specie facciamo alcuni esempi.
Orrorin tugenensis, come rilevato nello studio di Almécija et al. 2013, si situa in una posizione intermedia fra le scimmie mioceniche e le australopitecine, e potrebbe essere uno dei primissimi rappresentanti di Hominina, la sottotribù che proprio circa 6 mln di anni fa si separò dalla tribù Hominini, formando due rami, uno appunto quello di Hominina dalla quale discende H. sapiens (H. è l’abbreviazione di Homo) e l’altro, Panina, dalla quale discendono gli attuali scimpanzé e bonobo.
Oggi, il più antico Sahelanthropus tchadensis è sostanzialmente escluso dal ramo Hominina in quanto vissuto circa tra 7-6 milioni di anni fa, quindi oltre a quanto si è ricavato dal cosiddetto ”orologio molecolare”, attraverso il quale si è potuto stabilire una datazione di massima della separazione dal progenitore comune dei due rami Hominina e Panina a circa 6 mln di anni fa.
Per alcune datazioni è stato inserito il punto interrogativo in quanto certune classificazioni sono a mio avviso assai discutibili ed eventuali rivalutazioni potrebbero cambiare alcune cronologie.
Ad esempio, non è chiaro quando si sia estinto il genere Australopithecus. Gli ultimi esemplari certi sono quelli di Au. sediba datati a circa 1,95 mln di anni fa. Gli ultimi parantropi emersi dal registro fossile sono datati a circa 1,4-1,2 mln di anni fa ma, pur facendo parte delle australopitecine, non sono membri di Australopithecus in senso stretto.
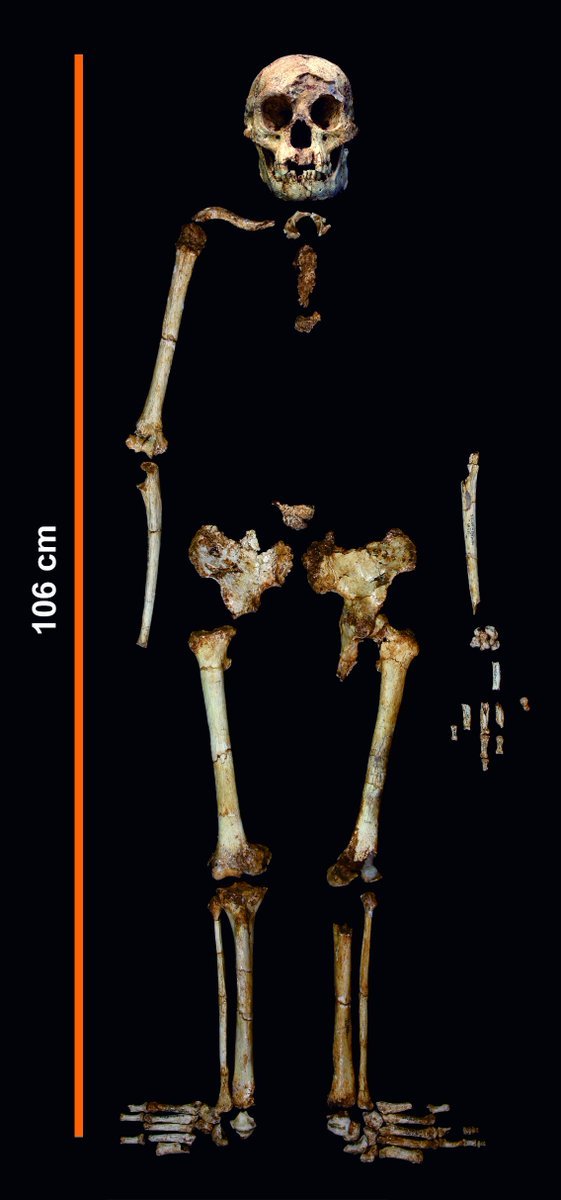
Homo Floresiensis
Tuttavia ci sono alcuni casi piuttosto dubbi, come H. naledi o H. floresiensis, che potrebbero mettere in discussione la data di estinzione di Australopithecus. Queste due specie presentano una morfologia cosiddetta a mosaico piuttosto particolare e anomala. Praticamente tutti gli ominini presentano generalmente un mosaico di caratteri, ovvero caratteri arcaici associati a caratteri nuovi (derivati), tuttavia le valutazioni vanno fatte a seconda della datazione. Un conto se si sta analizzando un ominino di 3 mln di anni fa, un conto se è di 300.000 anni fa.
Homo naledi è stato datato circa tra 235-336.000 anni fa, tuttavia presenta un cervello con dimensioni (circa 460-560cc) che rientrano nella gamma degli australopitechi/parantropi e addirittura in quella degli scimpanzé, a parte un singolo individuo che eccede appena leggermente di un 9% (610cc circa) e che potrebbe rientrare in una gamma di variazione individuale.
Tanto per dare un’idea, H. sapiens ha un cervello mediamente di 1.450cc per i maschi e 1.350cc per le femmine, ma a livello individuale le dimensioni del nostro cervello variano tra 910cc e ben 2.200cc!
Teniamo a mente che H. naledi è stato scoperto nel 2013 e gli studi sono stati pubblicati a partire dal 2015. Il Prof. Damiano Marchi, paleoantropologo che ha partecipato alle analisi dei resti di H. naledi, nel suo libro ”Il mistero di Homo naledi” (Mondadori, 2016) scrive:
Homo naledi
“Homo naledi associa una volta cranica e una mandibola dai tratti moderni a un cervello molto piccolo, con un volume che varia da 465 a 560 cc, paragonabile a quello dei rappresentanti di specie arcaiche come Australopithecus e Paranthropus. Peculiarità, questa, che ci fornisce una prova ulteriore del fatto che l’aumento delle dimensioni del cervello non costituisce una caratteristica del genere Homo presente fin dalle fasi iniziali della sua evoluzione, come ritenevano gli antropologi in passato…”

e prosegue affermando che:
“…va indiscutibilmente inserito nel genere Homo”.
 Dall’affermazione del Prof. Marchi la conclusione è ovvia: è Homo e in più ha un cervello piccolo, allora non può dare altro risultato che l’encefalizzazione non è più un carattere peculiare di Homo. La genetica però dice e dimostra l’esatto opposto.
Dall’affermazione del Prof. Marchi la conclusione è ovvia: è Homo e in più ha un cervello piccolo, allora non può dare altro risultato che l’encefalizzazione non è più un carattere peculiare di Homo. La genetica però dice e dimostra l’esatto opposto.
Ad oggi, purtroppo, in nessuno degli studi scientifici su H. naledi vi è il benché minimo riferimento agli studi genetici che sono stati pubblicati già diversi anni prima del ritrovamento del 2013. C’è da sottolineare che purtroppo non è stato nemmeno ancora trovato DNA di H. naledi, per cui la prova certa si avrà appena si riuscirà a trovarne.
Negli ultimi anni sono stati scoperti geni e regioni regolative del genoma umano che hanno determinato lo sviluppo del cervello, sia per quanto riguarda le dimensioni in generale che, soprattutto, della neocorteccia e della densità dei neuroni. Queste regioni regolative umano-specifiche sono state denominate “HAR”, Human Accelerated Region (Regioni Umane Accelerate).
Più specificatamente, per quanto riguarda l’encefalizzazione (crescita volumetrica e sviluppo dei processi inerenti alle funzioni organiche e psicologiche del cervello), sono di particolare importanza le regioni denominate HAR1 e HAR5, dove è stato riscontrato un tasso eccezionale di mutazioni positive totalmente assente nelle scimmie antropomorfe e in altri organismi, come è stato rilevato in diversi studi come quello di Pollard et al. del 2006 che riguarda la regione regolativa denominata HAR1.
In un articolo del 20 febbraio 2015 apparso sulla rivista online Le scienze.it dal titolo Un frammento di DNA fa grande il cervello dell’uomo (in relazione allo studio di Lomax Boyd et al. 2015), si legge:
“Una piccola sequenza di DNA non codificante del genoma umano è responsabile del grande sviluppo della corteccia cerebrale che caratterizza la nostra specie. Per quanto questa sequenza, chiamata HARE5, possa esplicare a pieno la sua funzione solo nell’essere umano, il suo inserimento in embrioni di topo ha fatto sì che i roditori sviluppassero una corteccia cerebrale di dimensioni inusualmente grande. Verso la fine della gestazione la differenza di dimensioni del cervello era diventata visibile a occhio nudo, alla nascita la neocorteccia dei topi con HARE5 umano era più grande del 12% rispetto ai topi con enhancer di scimpanzè.”
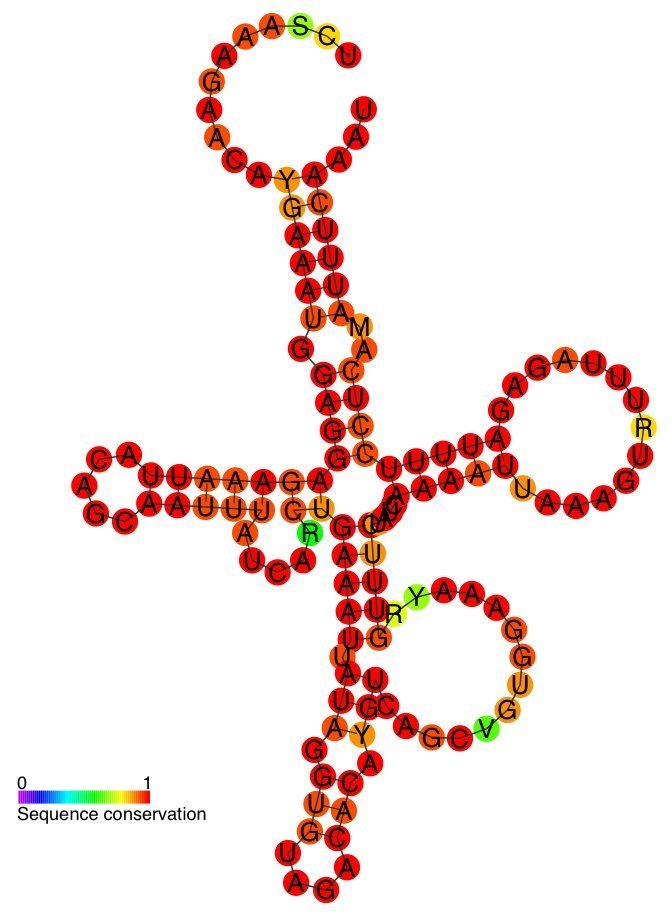
La struttura secondaria e la sequenza di conservazione di HAR1F
Di studi su geni che hanno determinato lo sviluppo del nostro cervello e, più specificatamente, della nostra corteccia cerebrale (neocorteccia), ne sono stati pubblicati diversi nel corso degli anni.
Nello studio di Florio et al., 2015, sul gene ARHGAP11B, in alcuni topi ai quali, attraverso tecniche genetiche, era stato trasferito il gene, fu osservato che la neocorteccia cresceva molto di più che nei topi normali non trattati.
Gli studi di Charrier et al. 2012 sul gene srGAP2, Suzuki et al. 2018 e Fiddes et al. 2018 sui geni NOTCH2NL, sono stati molto significativi non solo perché si è osservato quanto questi geni siano stati particolarmente influenti sullo sviluppo corticale e neuronale del nostro cervello, ma anche perché i primi gruppi sono apparsi tra i 3-4 mln di fa!!
Tutte queste regioni regolative e geni sono esclusivamente umane, per cui le implicazioni sono di conseguenza enormi. Ciò significa che è molto probabile che tutte le regioni regolative, e geni, esclusivamente umane che ad oggi sono state scoperte, in H. naledi non erano presenti, essendo vissuto in tempi così recenti (tra i 235-336.000 anni fa) e con un volume cranico ancora così particolarmente piccolo di appena 460-560cc (con un singolo caso di 610cc). Per cui se non possiede i geni che caratterizzano Homo è davvero strano che sia stato inserito nel genere Homo.
Se non bastasse, sono presenti in questo taxon anche tantissimi ulteriori caratteri morfologici tipicamente australopitecini. Non c’è solo l’aspetto genomico. È stato inserito un ulteriore punto interrogativo nella datazione del genere Homo e, più precisamente, riguardo alla sua emersione.
Fino a qualche tempo fa il più antico esemplare di Homo ritrovato risaliva a circa 2,4 mln di anni fa e attribuito al taxon (specie) Homo habilis. Tuttavia, una recente scoperta di una mandibola risalente a circa 2,8 mln di anni fa attribuita ad Homo (Villmoare et al., 2015) ha fatto decisamente retrodatare la presenza di Homo nel registro fossile (ricordo che le datazioni hanno una valenza solo relativa).
Questo fa capire che è sostanzialmente ancora ignota la data della comparsa di Homo, ovvero a quanto tempo risale la sua separazione dal progenitore comune condivisa, molto probabilmente, con almeno una delle varie specie di Australopithecus.

Omero distale di Australopithecus anamensis
Personalmente ipotizzo che il progenitore comune sia Au. anamensis, vissuto circa tra 4,2-3,5 mln di anni fa, e che l’altro ramo che si è originato da questa separazione sia Au. afarensis, il tutto avvenuto presumibilmente attorno a… 3,7 mln di anni fa!
Ci sono diversi indizi che portano a questa datazione sulla probabile emersione di Homo (questa volta in senso assolutivo e non solo relativo): i geni NOTCH2NL il cui primo gruppo ”A” è comparso proprio tra i 3-4 mln di anni fa, il gene srGAP la cui prima duplicazione è avvenuta circa 3,5 mln di anni fa (le datazioni genetiche non sono estremamente precise per cui c’è un certo margine di approssimazione), le impronte di Laetoli datate a 3,66 mln di anni fa le quali presentano caratteristiche morfologiche umane, un esemplare fossile denominato KSD-VP 1/1 Kadanuumuu, stranamente attribuito ad Au. afarensis, che presenta caratteri già umani come una gabbia toracica a ”campana”, diversamente da quella degli australopitechi che invece è generalmente a ”cono” o ”imbuto rovesciato” (per la cronaca H. naledi ha una gabbia toracica conica…).
A sostegno di questa ipotesi ci sono dunque robusti indizi sia genetici che paleontologici. Su H. naledi e su Kadanuumuu ci tornerò in futuro con articoli più approfonditi.
Concludendo, quello che si evince è che la nostra evoluzione è stata decisamente più variegata rispetto a come si credeva un tempo. Non è stata lineare, come se fosse avvenuta su una singola ”catena” dove si dovevano cercare anelli mancanti, ma è stata un ”cespuglio” ben più articolato. C’è ancora tantissimo da studiare e, si spera, da scoprire (e a mio modestissimo avviso anche da rivalutare…).
Di sicuro la nostra storia non finisce qui…
Autore: Alessandro Mori
Bibliografia:
– Buffa, Pietro I geni manipolati di Adamo (Uno editori)
– Marchi, Damiano Il mistero di Homo naledi (Mondadori)
– Tattersall, Ian I signori del pianeta (Codice edizioni)
– L’evoluzione del cervello (National Geographic)
– Corteccia cerebrale, migliaia di regolazioni geniche ci rendono umani (Le scienze.it, 2015)
– Il gene che controlla l’espansione della neocorteccia (Le scienze.it, 2015)
– Divisi dal DNA: il difficile rapporto tra archeologia e genomica (Le scienze.it, 2018)
– Più antica di 500.000 anni la comparsa del genere Homo (Le scienze.it, 2015)
– Un frammento di DNA fa grande il cervello dell’uomo (Le scienze.it, 2015)
– Un gene (mal) duplicato ha diretto l’evoluzione del cervello umano (Le scienze.it, 2012)
– Almécija et al., 2013 The femur of Orrorin tugenensis exhibits morphometric affinities with both Miocene apes and later hominins (doi: https://doi.org/10.1038/ncomms3888)
– Charrier et al., 2012 Inhibition of SRGAP2 function by its human-specific paralogs induces neoteny during spine maturation (doi: https://doi.org/10.1016/j.cell.2012.03.034)
– Fiddes et al., 2018 Human-specific NOTCH2NL genes affect notch signaling and cortical neurogenesis (doi: https://doi.org/10.1016/j.cell.2018.03.051)
– Florio et al., 2015, Human-specific gene ARHGAP11B promotes basal progenitor amplification and neocortex expansion (doi: https://doi.org/10.1126/science.aaa1975)
– Florio et al., 2016 A single splice site mutation in human-specific ARHGAP11B causes basal progenitor amplification (doi: https://doi.org/10.1126/sciadv.1601941)
– Laird et al., 2017 The skull of Homo naledi (doi: https://doi.org/10.1016/j.jhevol.2016.09.0099
– Lomax Boyd et al., 2015 Human-chimpanzee differences in a FZD8 enhancer alter cell-cycle dynamics in the developing neocortex (doi: https://doi.org/10.1016/j.cub.2015.01.041)
– Pollard et al., 2006, An RNA gene expressed during cortical development evolved rapidly in humans (doi: https://doi.org/10.1038/nature05113)
– Pollard et al., 2006, Forces shaping the fastest evolving regions in the human genome (doi: https://doi.org/10.1371/journal.pgen.0020168)
– Suzuki et al., 2018, Human-specific NOTCH2NL genes expand cortical neurogenesis through Delta/Notch regulation (doi: https://doi.org/10.1016/j.cell.2018.03.067)
– Villmoare et al., 2015 Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia (doi: https://doi.org/10.1126/science.aaa1343)



